Автор: Ю.В. Ковалева
Жировая ткань в роли эндокринного «органа

Жировая ткань продуцирует адипокины, влияющие на потребление пищи, метаболические процессы, формирование оксидативного стресса и нарушений функции сердечно-сосудистой системы [1-4]. Адипокины действуют эндо-, пара- и интракринным путем. Клеточное развитие и экспрессия генов в процессе дифференцировки адипоцитов представлены на рисунке 1.

Рис. 1. Клеточное развитие и экспрессия генов в процессе дифференцировки адипоцитов [по A.Shaffler и соавт. (2006) с модификацией]
Примечание. LPL-липопротеинлипаза; ИФР-1 – инсулиноподобный фактор роста 1-го типа; Krox20 – проадипогенный фактор; C/EBP, Ccaat-enhancer-binding proteins – Ccaat-энхансер-связывающий протеин; ADD1, adipocyte determination differentiation factor 1 – адипоцитарный, от детерминации- и от дифференцирования-зависимый фактор 1; PPARɣ, peroxisome proliferator-activated receptors ɣ - рецептор, активируемый пролифератором пероксисом ɣ; KLF – транскрипционные Kruppel-подобные факторы; GLUT-4 – внутриклеточный переносчик глюкозы; CORS-2G – коллагенозные повторы, содержащие последовательность 26 кДа протеина.
Продукты секреции адипоцитов, влияющие на развитие инсулинорезистентности и метаболических нарушений, приведены в таблице 1.
Таблица 1. Продукты секреции адипоцитов

Часть этих белков – провоспалительные цитокины, часть принимает участие в метаболизме липидов, в то время как другие вовлечены в работу системы комплемента и сосудистый гемостаз.
Лептин – секретируется адипоцитами пропорционально массе жировой ткани. Молекулярная масса белка - 16 кДа. В составе - 167 аминокислот; он имеет замкнутую внутримолекулярную дисульфидную связь, в крови циркулирует в виде мономера [5, 6].

Действие лептина основано на активации специфического лептинового рецептора, представленного длинной изоформой и многочисленными короткими. Рецепторы располагаются в гипоталамусе, мозжечке, коре, гиппокампе, сосудистых сплетениях и эндотелии мозговых капилляров. К настоящему времени клонировано по крайней мере 6 изоформ рецептора к лептину, с помощью которых опосредуются биологические влияния гормона.
Большинство известных эффектов лептина опосредуется через рецепторы LRb (leptin receptor b) в организме. Под действием лептина в клетках гипоталамуса усиливается транскрипция около 80 генов. Связывание лептина со специфическими рецепторами в гипоталамусе изменяет экспрессию ряда нейропептидов, регулирующих нейроэндокринную функцию, потребление и расход энергии в организме. Среди них наиболее изучены - нейропептид Y (NPY), тиреотропин-рилизинг- гормон и кортикотропин-рилизинг-гормон.
Установлено: уровень лептина в крови повышается при увеличении массы жировой ткани, причем его продукция в подкожной жировой клетчатке выше, чем в висцеральных жировых депо. Но концентрация лептина широко варьирует у людей с одинаковым показателем индекса массы тела (ИМТ), что подтверждает участие как генетических факторов, так и факторов окружающей среды в регуляции уровня этого гормона.
Концентрация лептина отражает не только количество накопленного жира, но и нарушение энергетического обмена. При продолжительном голодании отмечено значительное его снижение, при переедании – повышение. При этом состав питательных компонентов в рационе на него не влияет. Вместе с тем доказано, что при увеличении массы тела на 10% уровень лептина повышается на 300%, а при последующем снижении массы тела на 10% - снижается всего лишь на 53 %.
Уровень лептина в крови практически сразу же изменяется при любых колебаниях концентрации кортикотропина и кортизола у здоровых мужчин, лютеинизирующего гормона (ЛГ) и эстрадиола у здоровых женщин.
Экспрессия лептинового рецептора выявлена в периферических тканях - легких, почках, печени, поджелудочной железе, надпочечниках, яичниках, стволовых клетках гемопоэза и скелетных мышцах.
Дефицит лептина – вовсе не основная причина развития ожирения. Напротив, у лиц с ожирением очень часто отмечается повышенный уровень лептина.
Уровень лептина в сыворотке крови повышается с увеличением массы тела и степени ожирения, тогда как доказанная недостаточность секреции лептина встречается редко. Эти данные позволяют считать, что при ожирении имеет место резистентность к лептину на уровне транспорта в ЦНС или на пострецепторном уровне. Избыток лептина приводит к подавлению секреции инсулина и вызывает инсулинорезистентность скелетных мышц и жировой ткани.
Лептин, подобно инсулину, регулирует гомеостаз жирных кислот, предохраняя от развития липотоксикоза. В условиях лептинорезистентности происходят нарушения компенсаторного окисления избытка свободных жирных кислот и активация неокислительного пути их метаболизма. Это приводит к накоплению неокисленных продуктов метаболизма свободных жирных кислот и керамидов, нарушающих инсулинозависимое поглощение глюкозы тканями. В результате триглицериды откладываются как между волокнами скелетных и сердечной мышц, так и в клетках печени и миоцитах.
Таким образом, при избыточном развитии жировой ткани нарушение действия лептина может быть одним из ведущих факторов развития инсулинорезистентноети и нарушения функции β-клеток поджелудочной железы [1,9,10]. Возможная роль лептина в патогенезе сахарного диабета в настоящее время активно изучается. Высказывается предположение, что высокий уровень лептина может тормозить действие инсулина в печени, снижая активность ряда ферментов, регулирующих скорость глюконеогенеза, а в жировой ткани тормозит стимулированный инсулином транспорт глюкозы.
Обнаружено, что плазменный уровень лептина повышен при АГ, ХСН, ИМ. Как по данным экспериментальных работ in vitro, так и по результатам популяционных перекрестных и когортных исследований, лептин играет протективную роль в развитии гипертрофии левого желудочка.
Показано: дефицит лептина, связанный с наличием мутации гена лептина или лептинового рецептора, приводит к нарушениям функции гипоталамо-гипофизарно-гонадной системы и щитовидной железы.
В последнее время получены данные, что лептин секретируется плацентой и яичниками [12]. Рецепторы к лептину обнаружены на поверхности гранулезных клеток, клеток теки и интерстициальных клеток яичников. Установлено: лептин подавляет процессы стероидогенеза в гранулезных клетках и клетках теки, проявляя антагонизм по отношению к инсулиноподобному фактору роста 1 (ИФР-1), инсулину, ЛГ и трансформирующему фактору роста бета (ТФР-β). Кроме того, независимо от влияния на механизмы стероидогенеза высокие концентрации лептина подавляют развитие доминантного фолликула и нарушают овуляцию [6]. Есть данные о роли повышенного уровня лептина в патогенезе синдрома поликистозных яичников (СПКЯ). По результатам исследования Р.R. Brzechffa и соавт. (1996), у значительной части женщин с СПКЯ уровни лептина оказались выше, чем ожидалось исходя из показателей ИМТ, свободного тестостерона, чувствительности к инсулину.
Высказано предположение, что лептин меньше эволюционировал как гормон сытости и больше – как сигнал для репродуктивной системы об адекватном запасе энергии. Уровень лептина повышается на 50% непосредственно перед началом полового созревания, в дальнейшем наблюдается его стабилизация на протяжении 2 лет. Для девочек характерен стабильный подъем уровня лептина на протяжении всего пубертатного периода с более высокими показателями на поздних стадиях, что соотносится с увеличением эстрогенов. Существует критический уровень лептина, необходимый для запуска репродуктивной системы. При низком уровне лептина может развиться аменорея, что указывает на способность женского организма останавливать процесс овуляции при снижении массы жировой ткани, то есть недостаточности энергетических резервов.
В течение короткого времени в организме человека лептин может действовать как диуретический фактор, способствующий выведению натрия и задержке калия. Однако при длительном воздействии лептин стимулирует метаболизм норадреналина и повышает тонус симпатической нервной системы. Поэтому активно изучается роль лептина в патогенезе АГ.
1995 – 96 гг. – независимые группы японских и американских ученых открывают белковый гормон, синтезируемый адипоцитами, - адипонектин, название которого имеет многочисленные синонимы: ACRp30 (adipocyte complement-related protein 30 kDa), adipoQ и GBP28 (gelatin binding protein 28 kDa).
Адипонектин - продукт гена ожирения арМ 1 (adipose most abundant gene transcript 1), который локализуется на хромосоме 3q27. Это коллагеноподобный белок, представляющий собой полипептид с молекулярной массой 30 кДа, содержащий 244 аминокислотных остатка, сходный по структуре с молекулой коллагена и фактора некроза опухолей бета (ФНО-β) и циркулирующий в периферическом кровообращении в 8 различных изоформах. Адипонектин обладает противовоспалительным и антиатерогенным действием.
Концентрация адипонектина в крови обратно коррелирует с массой жировой ткани, т.е. его уровень при ожирении ниже, чем у людей с нормальной массой тела.
Многоцентровые исследования показали: уровень адипонектина плазмы обратно пропорционален степени ожирения, массе жировой ткани, отношению окружности талии к окружности бедер, дислипидемии и инсулинорезистентности [14-17]. Концентрация адипонектина в плазме крови ниже у женщин с высоким ИМТ (> 30 кг/м2), гипертриглицеридемией и гиперинсулинемией; вместе с тем уровень адипонектина у женщин с сахарным диабетом выше, чем у мужчин [17]. Выявлена положительная корреляция уровня адипонектина с возрастом [17].
В ряде исследований показано протективное действие адипонектина на развитие инсулинорезистентности. Он повышает чувствительность к инсулину путем активации липолиза, приводящей к снижению уровня циркулирующих жирных кислот и поступления их в печень, уменьшению содержания триглицеридов в печени и мышцах, и стимулирует окисление жирных кислот путем активации протеинкиназы, способствуя снижению продукции глюкозы печенью, синтеза триглицеридов, липопротеинов очень низкой плотности [18, 19]. Также способствует уменьшению инсулинорезистентности, стимулируя фосфорилирование тирозина рецептора инсулина. В мышечной ткани адипонектин стимулирует окисление свободных жирных кислот, уменьшает интрамиоцеллюлярные накопления липидов и улучшает чувствительность мышечной ткани к инсулину.
При ожирении, СД2 и ИБС уровень адипонектина в крови снижен [17,19]. Клинические исследования показали: низкий уровень адипонектина ассоциирован с атерогенным липидным профилем и высоким уровнем артериального давления. У пациентов с СД2 и ИБС уровень адипонектина ниже, чем у пациентов без ИБС, что может свидетельствовать о его антиатерогенных свойствах. Предполагается, что адипонектин обладает антиатерогенными эффектами за счет стимуляции окисления жирных кислот и снижения уровня триглицеридов в плазме. Обнаружена взаимосвязь между низким уровнем адипонектина, ожирением, инсулинорезистентнотью, ИБС и дислипидемией [21]. Содержание адипонектина в сыворотке крови обратно коррелирует с уровнем триглицеридов, атерогенным индексом, аполипопротеинами (Аро) АроВ или АроЕ и положительно – с липопротеинами высокой плотности и уровнем АроА-1.
Адипонектин рассматривается как кардиопротективный гормон, нормальное содержание которого ассоциировано со снижением оксидативного стресса, гипертрофии левого желудочка и воспаления [11].
Снижение уровня адипонектина может способствовать развитию связанных с ожирением метаболических нарушений и/или заболеваний, например, СПКЯ. Выявлено существенное снижение уровня адипонектина у женщин с СПКЯ, особенно при наличии ожирения, но сравнению с таковым у женщин с нормальной массой тела [16, 22, 23]. При этом низкий уровень адипонектина обусловлен именно иисулинорезистентностью, выявляемой у пациенток с СПКЯ, а не гиперандрогенией [23].
Адипонектин способен подавлять образование тестостерона в яичниках [2, 27].
Высвобождаясь в кровеносную систему, адипонектин накапливается в сосудистой стенке в ответ на повреждение эндотелия и модулирует воспалительный процесс в эндотелии. Противовоспалительные эффекты адипонектина включают ингибирование экспрессии адгезивных молекул в эндотелиальных клетках, снижение адгезии моноцитов, ингибирование образования цитокинов макрофагами [24], а также снижение экспрессии С-реактивного белка в жировой ткани [25]. Вследствие этого происходит подавление воспалительных процессов.
Помимо этого, адипонектин ингибирует ФНО-α-индуцируемую активацию нуклеарного фактора каппа би (NF-кВ) в эндотелиоцитах и гладкомышечных клетках аорты человека посредством ингибирования фосфорилирования 1кВ [25]. Физиологические концентрации адипонектина ингибируют сигнализацию NF-кВ в макрофагах и эндотелиальных клетках, что приводит к снижению экспрессии ФНО-α и молекул клеточной адгезии – молекулы адгезии сосудистого эндотелия 1 (vascular cell adhesion molecule 1, VCAM-1), молекулы межклеточной адгезии (intercellular adhesion molecule 1, ICAM-1) и Е-селектина, а также выработки антагонистов к рецепторам противовоспалительных медиаторов - интерлейкино (ИЛ) 10 и 1 — в лейкоцитах человека [24-26].
Адипсин - гормон, который играет важную роль в липолизе и поддержании нормального «адипостата». Обладает сериновой протеазной активностью, идентичной таковой у комплементарного фактора D, продукция которого является начальным шагом в активации альтернативного пути системы комплемента.
Повышение уровня адипсина в крови сопровождается потерей массы жировой ткани и увеличением скорости метаболических процессов.
Изменение секреции адипсина, по-видимому, играет важную роль в патогенезе ожирения. Так, при ожирении, индуцированном диетой, уровень адипсина повышен и положительно коррелирует с индексом массы тела.
Предполагают, что адипсин может влиять на образование триглицеридов опосредованно через генерацию синтеза белка, стимулирующего ацелирование.
2001 г. – выделен полипептид, названный резистином. Секретируется преимущественно преадипоцитами и в меньшей степени зрелыми адипоцитами в основном абдоминальной локализации [4, 28].
Резистин, или адипоцитспецифический секреторный фактор (ADSF/FIZZ3), - пептид, состоящий из 114 аминокислотных остатков, принадлежит к семейству цистеинсодержащих С-терминальных доменовых белков, называемых резистинподобными (resistinlike molecules, RELM) или FIZZ молекулами, вовлеченными в процессы воспаления. Ген резистина локализуется на хромосоме 19р13.3.
Резистин участвует в регуляции углеводного обмена как антагонист инсулина, нейтрализуя тормозящее влияние инсулина на продукцию глюкозы печенью и снижая поглощение глюкозы скелетной мускулатурой независимо от транспортера глюкозы (glucose transporter type 4) GLUT-4. Изучение биологического действия резистина, секретируемого адипоцитами и эндокринными клетками ЖКТ, показало: резистин индуцирует печеночную, но не периферическую резистентность к инсулину у крыс и, таким образом, отвечает за повышение скорости образования глюкозы печенью [29].
Убедительных данных о связи уровня резистина с ожирением и инсулинорезистентностью нет, хотя установлено, что при повышении резистина в крови инсулино- резистентность возрастает. Резистин снижает инсулинстимулированное поглощение глюкозы и может нарушать дифференцировку адипоцитов.
Есть данные, что резистин способен увеличивать синтез тестостерона в яичниках [2].
В период эмбрионального развития ген резистина экспрессируется в плаценте преимущественно в конце беременности, и его содержание в плазме крови беременных женщин значительно выше. Считается, что в этот период резистин выполняет роль регулятора углеводного обмена.
Установлено также, что повышенная экспрессия гена резистина в жировой ткани при центральном ожирении коррелирует с СД2 и сердечно-сосудистых заболеваний (ССЗ), что позволяет предположить возможность влияния этого гормона на связь СД и ССЗ с центральным ожирением [30].
Жировая ткань – место образования провоспалительных цитокинов – ФНО и некоторых интерлейкинов.
Медиаторы воспаления - патогенетический компонент осложнений ожирения, метаболической) синдрома, СД2, атеросклероза и ССЗ. Исследования, посвященные изучению роли провоспалительных цитокинов (ФНО-α и β, ИЛ-6, а также С-реактивного белка), позволили предположить, что воспаление участвует в патогенезе инсулинорезистентности, а указанные цитокины служат предикторами сосудистых осложнений сахарного диабета [31-34]. Биохимические маркеры воспаления (ФНО-α, 2-й тип растворимого рецептора ФНО-α, ИЛ-6, С-реактивный белок и растворимая молекула межклеточной адгезии 1-го типа) снижают чувствительность к инсулину, и жировая ткань полностью определяет взаимоотношения между цитокинами и инсулинорезистентностью.

Экспрессируется лимфоцитами и адипоцитами и обладает скорее ауто- и паракриннымн, чем эндокринными эффектами.
При ожирении имеет место повышенная экспрессия его гена как в жировой ткани, так и в мышцах [35], возрастает экспрессия рецептора 2-го типа к ФНО-α в жировой ткани. Уровень ФНО-α в жировой ткани коррелирует с массой жировой ткани и гипериисулинсмией. ФНО-α стимулирует секрецию лептина, и действие его опосредовано ИЛ-1. Введение ФНО-α вызывает уменьшение потребления пиши, задержку опорожнения желудка, ингибирует действие инсулина, модулирует уровни глюкагона и глюкокортикоидов, стимулирует термогенез.
Вследствие повышенной секреции ФНО-α и ИЛ-6 висцеральный жир оказывает провоспалительный эффект. Данные провоспалительные цитокины активируют фактор транскрипции ответа на окислительный стресс NF-kB [36].
ФНО-α сегодня рассматривается в качестве основного медиатора инсулинорезистентности в жировой ткани. Избыточное содержание жировой ткани ассоциировано с повышением продукции ФНО-α, что индуцирует развитие инсулинорезистентности, при этом уровень ФНО-α коррелирует с ее степенью и тяжестью [34].
Механизм действия ФНО-α на чувствительность к инсулину - снижение активности тирозинкиназы инсулинового рецептора и усиление фосфорилирования серина IRS-1 – субстрата инсулинового рецептора, торможениt экспрессии внутриклеточных переносчиков глюкозы (GLUT-4) в жировой и мышечной ткани. Кроме того, ФНО-α может уменьшать сигнал непосредственно через рецепторы, активируемые пероксисомными пролифераторами гамма (peroxisome proliferator- activated receptors у, PPAR-y). ФНО-а также может способствовать развитию инсулинорезистентности непрямым путем, через стимуляцию липолиза в адипоцитах. При снижении массы тела уровень ФНО-α снижается, и чувствительность к инсулину возрастает [40].
Цитокин ФНО-β имеет молекулярную массу 17 кДа и экспрессируется как в иммунных, так и в неиммунных клетках, включая эндотелиоциты, фибробласты и адипоциты.
Повышение содержания ФНО-β в сыворотке крови сочетается с наличием ожирения, инсулинорезистентностью, увеличением концентрации С-реактивного белка и ИЛ-6, ускорением апоптоза [36].
Установлено: высвобождение ФНО-β из клеток жировой ткани аналогично его высвобождению из моноцитов или макрофагов. Повышение экспрессии гена ФНО-β в адипоцитах животных и при ожирении у человека сопровождается повышением выраженности инсулинорезистентности. Значит, данный цитокин – один из ключевых медиаторов ее развития. Антиинсулиновое действие ФНО-β – следствие его влияния на снижение экспрессии GLUT-4 и ингибирования тирозинкиназы рецепторов инсулина в клетках и тканях-мишенях.
Пропорционально увеличению массы жировой ткани в крови нарастает концентрация ИЛ-6 [34]. Жировые клетки сальника продуцируют в 2-3 раза больше ИЛ-6, чем адипоциты подкожной локализации. В связи со спецификой расположения висцеральной жировой ткани секретируемый ею ИЛ-6 имеет возможность прямого воздействия на метаболические процессы путем подавления чувствительности рецепторов инсулина непосредственно в печени.
Уровень экспрессии гена ИЛ-6 в жировой ткани напрямую коррелирует как со степенью активации поглощения глюкозы, так и с выраженностью инсулинорезистентности in vivo и in vitro [34, 40]. Внутривенное введение ИЛ-6 у человека сопровождается повышением уровня свободных жирных кислот и глицерола в сыворотке крови, что является следствием его влияния на липолиз жировой ткани.
По современным представлениям, ИЛ-6 – вещество, чувствительное к уровню глюкозы, способствует снижению уровня гликогена мышц, стимулирует выработку глюкозы в печени и улучшает поглощение глюкозы скелетной мускулатурой. При выполнении физических нагрузок при истощении запасов гликогена уровень ИЛ-6 существенно повышается. У пациентов с СД2 базальный уровень ИЛ-6 в плазме более высок.
БСА (acylation stimulating protein, ASP), или C3adesArg - белок, стимулирующий ацетилирование.
Считается, что БСА образуется в результате взаимодействия факторов комплемента: СЗ, В и D (адипсин). Показано, что липопротеины (в частности, хиломикроны) влияют на увеличение высвобождения БСА.
Содержание БСА в сыворотке крови человека обратно связано с распределением глюкозы в организме в условиях эугликемической клэмп-методики [42]. Это может свидетельствовать о его роли в формировании чувствительности тканей к инсулину. БСА вовлечен в обмен жиров (ингибирует гормон-чувствительную липазу, повышает активность диацилглицеринтрансферазы и этерификацию жирных кислот, синтез триглицеридов, увеличивая их депонирование в жировых депо, снижает липолиз и выделение неэтерифицированных жирных кислот из адипоцитов) и углеводов (увеличивает поглощение глюкозы периферическими тканями и повышает усвоение глюкозы адипоцитами путем стимуляции транслокации транспортера глюкозы к периферии клетки, а также усиления глюкозостимулированной секреции инсулина В-клетками поджелудочной железы) [43]. Хотя эти эффекты проявляются независимо, они дополняют действие инсулина. Концентрация БСА в сыворотке крови повышена у больных с ожирением, инсулинорезистентностью, дислипидемией, СД2 и ИБС.

Рис. 2. Гормоны жировой ткани: мишени, регуляция, взаимодействия
Примечание: БСА – белок,стимулирующий ацетилирование; ЦНС- центральная нервная система;м СЖК – свободные жирные кислоты; ЛПВП- липопротеины высокой плотности; ЛПОНП – липопротеины очень низкой плотности; АМРК – аденозин монофосфат-киназа; ФНО-α – фактор некроза опухоли α; ИЛ-6 – интерлейкин-6.
Висфатин. Открыт в 2005 г. [4]. Представляет собой белок массой 52 кДа, синтезируемый висцеральными адипоцитами. Является острофазовым белком воспаления и повышается при синдроме острого поражения легких. Висфатин угнетает развитие апоптоза, который был активирован нейтрофилами. Играет физиологическую роль в регуляции инсулинпродуцирующих клеток [44] и метаболизма мышечных клеток [45].
Показано: эффекты висфатина по накоплению жировых депо реализуются через инсулиновые рецепторы. Связываясь с последними, висфатин их активирует. Введение рекомбинантного висфатина у мышей действует на инсулиновый рецептор аналогично инсулину [46]. Уровень висфатина в циркулирующих клетках крови напрямую коррелирует с ИМТ, окружностью талии и индексом инсулинорезистентности. Его уровень повышен при ожирении [47] и СД2 [48]; при этом он отрицательно коррелирует с маркерами воспаления и выраженностью артериосклероза [47, 49].
В многоядерных клетках крови у больных СД2 количество митохондриальной рибонуклеиновой кислоты (мРНК) висфатина в несколько раз выше по сравнению с пациентами с СД и дефицитом массы тела, или практически здоровыми лицами. Считается, что висфатин участвует в атерогенезе, в патогенезе АГ при ожирении и сосудистых осложнений сахарного диабета. Уже сейчас понятно, что висфатин - важный иммунорегулятор с выраженными противовоспалительными свойствами.
Васпин (visceral adipose tissue-derived serpin, серпин A12) – член семейства ингибиторов сериновых протеаз с массой 45-50 кДа, который синтезируется адипоцитами висцеральной жировой ткани [50-52].
Комплементарная дезоксирибонуклеиновая кислота (кДНК) васпина была изолирована из висцеральной белой жировой ткани крыс с ожирением линии Otsuka Long-Evans Tokushima fatty (OLETF) - животной модели абдоминального ожирения при сахарном диабете 2-го типа [50, 51].
Человеческий, мышиный и крысиный васпины состоят из 395, 394 и 392 аминокислотных остатков соответственно, этот адипокин имеет около 40% гомологии с 1-антитрипсином [50].
Васпин относится к группе серпинов, состоящей из неоднородных по своей структуре молекул, угнетающих протеазную активность. Кроме того, серпины принимают участие в метаболизме и транспорте гормонов, угнетении воспалительного каскада, регуляции свертывания крови и ангиогенезе. У человека васпин синтезируется не только висцеральными, но и подкожножировыми адипоцитами. Обнаружена секреция васпина в коже, гипоталамусе, печени, островках поджелудочной железы, желудке [51, 53].
Экспрессия васпина тесно коррелирует с количеством жировой ткани и достаточно редко обнаруживается у худых людей. Так, секреция мРНК васпина не определялась у пациентов с нормальной массой тела (индекс массы тела < 25 кг/м2) и наиболее часто определялась у пациентов с СД2 [54].
Экспрессия висцерального васпина значительно коррелирует с ИМТ, процентом жира и концентрацией глюкозы в плазме крови через 2 часа при проведении глюкозотолерантного теста.
Экспрессия мРНК васпина в подкожной жировой ткани также коррелирует с инсулином плазмы крови натощак и скоростью инфузии глюкозы при проведении эугликемического клэмп-теста [54]. Низкое содержание васпина в крови ассоциировано с высоким уровнем физической активности [52]. Так, сывороточные концентрации васпина, как было показано, ниже у худощавых субъектов и спортсменов с длительным опытом физической активности [55]. В то же время физическая активность у нетренированных людей вызывает повышение уровня васпина: курс физических упражнений в течение 4 недель сопровождается существенным повышением концентрации этого адипокина в крови [52, 55].
Уровень васпина у женщин значительно выше, чем у мужчин [52]. При этом гендерные различия в концентрации васпина формируются еще в пубертатном периоде: у девочек его содержание в крови выше, чем у мальчиков [53].
Обнаружено, что у девочек концентрация васпина повышается с возрастом и стадией пубертата, тогда как у мальчиков таких изменений не наблюдается [53].
Секреция васпина повышена при ожирении, инсулинорезистентности и СД2 [51-55]. Васпин уменьшает определенные отклонения, наблюдаемые при ожирении, ассоциированном с сахарным диабетом, и метаболическом синдроме, повышая толерантность к глюкозе и чувствительность тканей к инсулину, особенно в клетках белой жировой ткани, а также нормализуя экспрессию генов, активация которых способствует развитию инсулинорезистентности [50]. При назначении рекомбинантного человеческого васпина повышается чувствительность к инсулину и нормализуется экспрессия генов жировой ткани. Таким образом, васпин рассматривается как адипокин, обладающий способностью повышать чувствительность к инсулину [53-55].
Апелин секретируется адипоцитами и эндотелиальными клетками. Уровень повышен при ожирении, особенно в сочетании с гиперинсулинемией [56] и гиперхолестеринемией [57]. Апелин регулирует диаметр кровеносных сосудов при ангиогенезе [58]. Повышенная секреция апелина ассоциирована с воспалительной реакцией [59].
Секреция апелина угнетается при голодании и вновь увеличивается при последующем приеме пищи. Инсулин способен непосредственно регулировать секрецию апелина, по-видимому, путем контроля в адипоцитах экспрессии генов, ответственных за его синтез [56]. Тот факт, что инсулин способен контролировать секрецию адипокинов, по крайней мере, некоторых, указывает на весьма сложные пути влияния этого гормона на метаболизм, на наличие не только широко известных прямых, но и опосредованных эффектов.
Оментин синтезируется адипоцитами висцерального жира. Уровень повышен при ожирении и инсулинорезистентности [60]. Полагают, что физиологическое значение этого адипокина сводится к модуляции периферических эффектов инсулина [61,62].
Выявлено благоприятное влияние оментина на воспаление, гомеостаз глюкозы и протективный эффект в отношении сердечно-сосудистой патологии [63].
Выводы
Экспрессия адипокинов зависит от различных параметров: общая жировая масса, процентное содержание и характер распределения жира. Положительная корреляция с массой белой жировой ткани наблюдается у всех адипокинов, кроме ади- понектина и оментина [64].
Гормоны жировой ткани, за исключением адипонектина, снижают чувствительность периферических тканей к инсулину, что сопровождается повышением степени выраженности инсулинорезистентности (рис. 3).
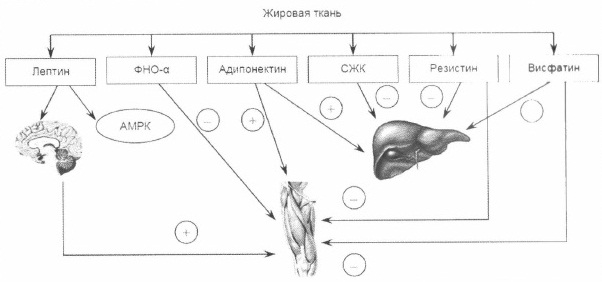
Рис. 3. Влияние гормонов жировой ткани на чувствительность к инсулину
Примечание. ФНО-α – фактор некроза опухоли α; СЖК – свободные жирные кислоты
АМРК – аденозин монофосфат-киназа.Большая выраженность инсулинорезистентности у лиц с абдоминальным типом ожирения обусловлена неодинаковой экспрессией генов гормонов жировой ткани в абдоминальной и подкожной жировой клетчатке (табл. 2).
Таблица 2. Различия в секреции гормонов подкожной и висцеральной жировой ткани

Как следует из таблицы, висцеральная жировая клетчатка секретирует значительно более высокое количество гормонов, усиливающих проявление инсулинорезистентности (резистин и другие), причем в висцеральной клетчатке снижается секреция гормонов, влияние которых заключается в снижении степени выраженности инсулинорезистентности (адипонектин).
Дисфункция жировой ткани характеризуется снижением чувствительности к инсулину, гипоксией, повышением параметров внутриклеточного стресса, повышением аутофагии и апоптоза, развитием воспаления [66]. В результате адипоциты запускают сигналы, способствующие возникновению провоспалительного, диабетогенного и атерогенного профиля сыворотки крови.
Жировая ткань – одно из важных мест активной продукции и метаболизма стероидов, что обеспечивается активностью ароматаз, позволяющих конвертировать фракции циркулирующих андрогенов (андростендион и тестостерон) в эстрогены (эстрон и эстрадиол соответственно).
Связь между количеством жировой ткани и содержанием эстрогенов выявлена в ряде исследований. Выраженность ароматизации существенно коррелирует с массой жира. Жировая ткань содержит и другие ферментные системы, такие как 17β-гидроксистероид- дегидрогеназа (17β-ГСД), которая катализирует трансформацию эстрадиола в эстрон, андростендиона в тестостерон, дегидроэпиандростерон-сульфат (ДЭА) в Δ4-андростендиол (А-диол).
Значительные количества стероидов сохраняются в жировой ткани. Так, положительный градиент ткань/плазма подтвержден для обоих эстрогенов и основных андрогенов (ДЭА, андростендион, тестостерон, А-диол), за исключением ДЭА-С. Как и в плазме, концентрация половых гормонов в жировой ткани с возрастом снижается.
Литература:
1. Bliiher M, Mantzoros CS. From leptin to other adipokines in health and disease: facts and expectations at the beginning of the 21st century. Metabolism. 2015;64( 1): 131—45. doi: 10.1016/j. metabol.2014.10.016
2. Mitchell M, Armstrong DT, Robker RT, Norman RJ. Adipokines: implications for female fertility and obesity. Reproduction. 2005; 130(5 ):583—597.
3. SchafflerA,Muller-LadnerU,Sch6lmerichJ,BuchlerC. Role of adipose tissue as an inflammatory organ in human diseases. EndocrRev. 2006;27(5):449-67.
4. Smitka K, Maresova D. Adipose tissue as an endocrine organ: an update on pro-inflammatory and anti-inflammatory microenvironment. Prague Med Rep. 2015; 116(2):87— 111. doi: 10.14712/23362936.2015.49.
5. Galic S, Oakhill JS, Steinberg GR. Adipose tissue as an endocrine organ. Mol Cell Endocrinol. 2010;316(2): 129—139.
6. 1 kinsman GJ, Barb CR, Lents CA. Leptin and reproductive function. Biochimie. 2012:94(101:2075 2081.
7. McMillen JC, Edwards LJ, Duffield J, Muhlhausler BS. Regulation of leptin synthesis and secretion before birth: implications for the early programming of adult obesity. Reproduction. 2006;131f3):415—427.
8. Shimizu H, Oh IS. Okada S. Mori M. Leptin resistance and obesity. Endocr J. 2007;54( 1): 17—26.
9. Antuna-Puente B, Feve B, Fellahi S, Bastard JP. Adipokines: the missing link between insulin resistance and obesity. Diabetes Metab. 2008;34( 1):2-11.
10. Bliiher M. Adipose tissue dysfunction in obesity. Exp Clin Endocrinol Diabetes. 2009; 117(6):241—250.
11. Ghantous CM, Azrak Z, Hanache S, Abou-Kheir W, Zeidan A. Differential role of leptin and adiponectin in cardiovascular system. Int J Endocrinol. 2015;2015:534320. doi: 10.1155/2015/534320
12. Hausman GJ, Barb CR. Adipose tissue and the reproductive axis: biological aspects. Endocr Dev. 2010; 19:31 —44.
13. Raucci R, Rusolo F, Sharma A, Colonna G, Castello G, Costantini S. Functional and structural features of adipokine family. Cytokine. 2013;61 (1): 1 -14. doi: 10.1016.
14. Guerre-Millo M. Adiponectin: an update. Diabetes Metab. 2008;34( 1): 12—8.
15. Matsuzawa Y. Adiponectin: a key player in obesity related disorders. Curr Pharm Des. 2010; 16( 17): 1896-1901.
16. Groth SW. Adiponectin and polycystic ovary syndrome. Biol Res Nurs. 2010; 12( 1 ):62-72. doi: 10.1177/1099800410371824
17. AleidiS,IssaA,BustanjiH, Khalil M.Bustanji Y. Adiponectin scrum levels correlate with insulin resistance in type 2 diabetic patients. Saudi Pharm J. 2015;23(3):250 256. doi: 10.101 G/j. jsps.2014.11.011
18. Tilg H, Moschen AR. Role of adiponectin and PBEF/visfatin as regulators of inflammation: involvement in obesity-associated diseases. Clin Sci (Lond). 2008; 114(4):275—288. doi: 10.1042/CS20070196
19. Spranger J, Kroke A, Mohlig M, Bergmann MM, Ristow M, Boeing H et al. Adiponectin and protection against type 2 diabetes mellitus. Lancet. 2003;361 (9353):226-228.
20. Park SE, Park CY, Sweeney G. Biomarkers of insulin sensitivity and insulin resistance: Past, present and future. Crit Rev Clin Lab Sci. 2015;4:1-11.
21. Okamoto Y, Arita Y, Nishida M, Muraguchi M, Ouchi N, Takahashi M et al. An adipocyte-derived plasma protein, adiponectin, adheres to injured vascular walls. Horm Metab Res. 2000;32(2):47-50. doi: 10.1055/s-2007-978586.
22. Orio F Jr, Palomba S, Cascella T, Milan G, Mioni R, Pagano C et al. Adiponectin levels in women with PCOS. J Clin Endocrinol Metab. 2003;88(6):2619-2623.
23. Toulis KA, Goulis DG, Farmakiotis D, Georgopoulos NA, Katsikis I, Tarlatzis BC et al. Adiponectin levels in women with polycystic ovary syndrome: a systematic review and a meta-analysis. Hum Reprod Update. 2009; 15(3):297—307. doi: 10.1093/humupd/dmp006
24. Yokota T, Oritani K., Takahashi I, Ishikawa J, Matsuyama A, Ouchi N et al. Adiponectin, a new member of the family of soluble defense collagens, negatively regulates the growth of myelomonocytic progenitors and the functions of macrophages. Blood. 2000;96(5): 1723-1732.
25. Ouchi N, Kihara S, Arita Y, Okamoto Y, Maeda K, Kuriyama H et al. Adiponectin, an adipocyte-derived plasma protein, inhibits endothelial NF-кВ signaling through a cAMP- dependent pathway. Circulation. 2000; 102(11): 1296-1301.
26. Wolf AM, Wolf D. Rumpold H, Enrich B, Tilg H. Adiponectin induces the anti-inflammatory cytokines IL-10 and IL- IRA in human leukocytes. Biochem Biophys Res Commun. 2004;323(2):630-635.
27. RiestraP, Garcia-Anguita A, Ortega L.Garces C. Relationship of adiponectin with sex hormone levels in adolescents. Horm Res Paediatr. 2013;79(2):83-87.
28. Fischer-Posovszky P, Wabitsch M, Hochbcrg Z. Endocrinology of adipose tissue — an update. Horm Metab Res. 2007;39(5):314-321.
29. Waki H, Tontonoz P. Endocrine functions of adipose tissue. Annu Rev Pathol. 2007;2:31-56.
30. Fonseca-Alaniz MH, Takada J, Alonso-Vale MI, Lima FB. Adipose tissue as an endocrine organ: from theory to practice. J Pediatr (Rio J). 2007;83(Suppl 5):S192-203.
31. Maury E, Brichard SM. Adipokine dysregulation, adipose tissue inflammation and metabolic syndrome. Mol Cell Endocrinol. 2010;314(1):1—16. doi: 10.1016/j.mce.2009.07.031
32. Bastard JP, Maachi M, Lagathu C, Kim MJ, Caron M, Vidal H et al. Recent advances in the relationship between obesity, inflammation, and insulin resistance. Eur Cytokine Netw. 2006; 17 (1):4-12.
33. Heilbronn LK, Campbell LV. Adipose tissue macrophages, low grade inflammation and insulin resistance in human obesity. Curr Pharm Des. 2008; 14( 12): 1225-1230.
34. Thomas S,SureshS,SudheeshM,VijayakumarT. Association of insulin resistance with adipocytokine levels in patients with metabolic syndrome, ndian J Clin Biochem. 2015;30(2):155—160. doi: 10.10077s 12291-014-0423-7
35. Клебанова E.M., Балаболкин М.И. Гормоны жировой ткани и их роль в патогенезе сахарного диабета 2-го типа. Лечащий врач. 2010; 11:27-33. [Klebanova ЕМ, Balabolkin MI. 1 lonnones of adipose tissue and their role in the pathogenesis of type 2 diabetes mellitus. Lechaschiy vrach = Physician. 2010; 11:27-33. In Russian).
36. Coppack SW Pro-inflammatory cytokines and adipose tissue. Proc Nutr Soc. 2001 ;60 (3):349—356.
37. Axclsson .1. Hcimbtirgcr O, Lindholm B, Stenvinkel P. Adipose tissue and its relation to inflammation: the role of adi- pokines. .1 Ren Nutr. 2005; 15(1): 131 136.
38. Calabro P, Limongelli G, Pacileo G, Di Salvo G, Golino P, Calabro R. The role of adiposity as a determinant of an inflammatory milieu. J Cardiovasc Med (Hagerstown). 2008;9(5):450-60.
39. Grant RW, Stephens JM. Fat in flames: Influence of cytokines and pattern recognition receptors on adipocyte lipolysis. Am J Physiol Endocrinol Metab. 2015: ajpendo.00053.2015. doi: 10.1 !52/ajpendo.00053.2015
40. Bastard JP, Jardel C. Elevated levels of interleukin 6 arc reduced in serum and subcutaneous adipose tissue of obese women after weight loss. J Clin Endocrinol Metab. 2000;85 (9):3338—3342.
41. Tsai S, Clemente-Casares X, Revelo XS, Winer S, Winer DA. Are obesity-related insulin resistance and type 2 diabetes autoimmune diseases? Diabetes. 2015;64(6): 1886—1897. doi: 10.2337/dbl 4—1488
42. Havel PJ. Update on adipocyte hormones. Regulation of energy balance and carbohydrate/lipid metabolism. Diabetes. 2004;53(Suppl 1):S143-151.
43. Cianflone K, Xia Z, Chen LY. Critical review of acylation- stimulating protein physiology in humans and rodens. Biochem Biophys Acta. 2003;1609(2):127-143.
44. Tanaka T, Nabescima Y. Nampt/PBEF/Visfatin: a new player in beta cell physiology and in metabolic diseases? Cell Metab. 2007;6(5):341—343.
45. Krzysik-Walker SM, Ocon-Grove OM, Maddineni SR, Hendricks GL. 3rd, Ramachandran R is visfatin an adipokine or tnyokine? Evidence for greater visfatin expression in skeletal muscle than visceral fat in chickens. Endocrinology. 2008; 149 (4): 1543-1550.
46. Sethi JK, Vidal-Puig A. Visfatin: the missing link between intra-abdominal obesity and diabetes? Trends Mol Med. 2005; 11 (8):344—347.
47. Araki S, Dobashi K, Kubo K, Kawagoe R, Yamamoto Y, Kawada Y et al. Plasma visfatin concentration as a surrogate marker for visceral fat accumulation in obese children. Obesity. 2008; 16 (2):384-348. doi: 10.1038/oby.2007.54
48. Algasham A A, Barakat YA. Serum visfatin and its relation to insulin resistance and inflammation in type 2 diabetic patients with and without macroangiopathy. Saudi Med J. 2008;29(2): 185-192.
49. Moschen AR, Kaser A, Enrich B, Mosheimer B;, Theurl M, Niederegger H et al. Visfatin, an adipocytokine with proinflammatory and imminomodulating properties. J Immunol. 2007; 178(3): 1748-1758.
50. Hida K, Wada J, Eguchi J, Zhang H, Baba M, Seida A et al. Visceral adipose tissue-derived serine protease inhibitor: a unique insulin-sensitizing adipocytokine in obesity. Proc. Natl. Acad. Sci. USA. 2005:102(30): 10610 10615. doi: 10.1073/pnas. 0504703102
51. Bliiher M. Vaspin in obesity and diabetes: pathophysiological and clinical significance. Endocrine. 2012;41 (2): 176—182. doi: 10.1007/s 12020-011-9572-0
52. Youn BS, Kloting N. Kratzsch J, Lee N, Park JW, Song ES et al. Serum vaspin concentrations in human obesity and type 2 diabetes. Diabetes. 2008;57(2):372—377.
53. Komer A, Neef M, Friebe D, Erbs S, Kratzsch J, Dittrich К et al. Vaspin is related to gender, puberty and deteriorating insulin sensivity in children. Int JObes. 2011;35(4):578-586. doi: 10.1038/ ijo.2010.196
54. Kloting N. Bemdt J, Kralisch S, Kovacs P. Fasshauer M, Schon MR et al. Vaspin gene expression in human adipose tissue: association with obesity and type 2 diabetes. Biochem Biophys Res Commun. 2006;339(1 ):430-436.
55. Li Q, Chen R, Moriya J, Yamakawa J, Surnino H, Kanda T et al. A novel adipocytokine, visceral adipose tissue-derived serine protease inhibitor (vaspin), and obesity. J Int Med Res. 2008;36 (4):625-629.
56. Boucher J, Masri B, Daviaud D, Gesta S, Guigne C, Mazzucotelli A et al. Apelin, a newly identified adipokine up- regulated by insulin and obesity. Endocrinology. 2005:146 (4): 1764-1771.
57. Tasci I, Dogru T, Naharci I, Erdem G, Yilmaz MI, Sonmez A et al. Plasma apelin is lower in patients with elevated LDL-cholesterol. Exp Clin Endocrinol Diabetes. 2007:115 (7):428-432.
58. Kidoya H, Ueno M, Yamada Y, Mochizuki N. Nakata M,; Yano T et al. Spatial and temporal role of the apelin/APJsastem in the calibre size regulation of blood vessels during angiogenesis. EMBO J. 2008;27(3):522-534. doi: 10.1038/sj.emboj.7601982
59. Garcia-Diaz D, Campion J, Milagro FI, Martinez JA. Adiposity dependent apelin gene expression: relationships with oxidative and inflammation markers. Mol Cell Biochem. 2007;305 (l-2):87-94.
60. de Souza Batista CM, Yang RZ, Lee MJ, Glynn NM. Yu DZ, Pray J et al. Omentin plasma level and gene expression and are decreased in obesity. Diabetes. 2007;56(6):1655-1661.
61. Yang RZ, Lee MJ, Hu H, Pray J, Wu HB, Hansen BC et al. Identification of omentin as a novel depot-specific adipokine in human adipose tissue: possible role in modulating insulin action. Am J Physiol Endocrinol Metab. 2006;290(6):E1253-1261.
62. Wurrn S, Neumeier M, Weigert J, Schaffler A, Buechler C. Plasma levels of leptin, omentin, collagenous repeat- containing sequence of 26-kDa protein (COPS-26) and adiponectin before and after oral glucose uptake in slim adults. Cardiovasc Diabetol. 2007:6:7. doi: 10.1186/1475-2840-6-7
63. Tan YL, Zheng XL, Tang CK. The protective functions of omentin in cardiovascular diseases. Clin Chim Acta. 2015; pii: S0009-8981 (15)00282-X. doi: 10.1016/j.cca.2015.05.019
64. Fietta P, Delsante G. Focus on adipokines. Theor Biol Forum. 2013; 106(1-2): 103-29.
65. Matsuzawa Y. The role of fat topology in the risk of disease. Int J Obes (Lond). 2008;32(Suppl 7):S83-92.
66. Kloting N, Blither M. Adipocyte dysfunction, inflammation and metabolic syndrome. Rev Endocr Metab Disord. 2014; 15 (4):277—287. doi: 10.1007/sl 1154-014—930
Источник: журнал «Артериальная гипертензия», № 4, 2015